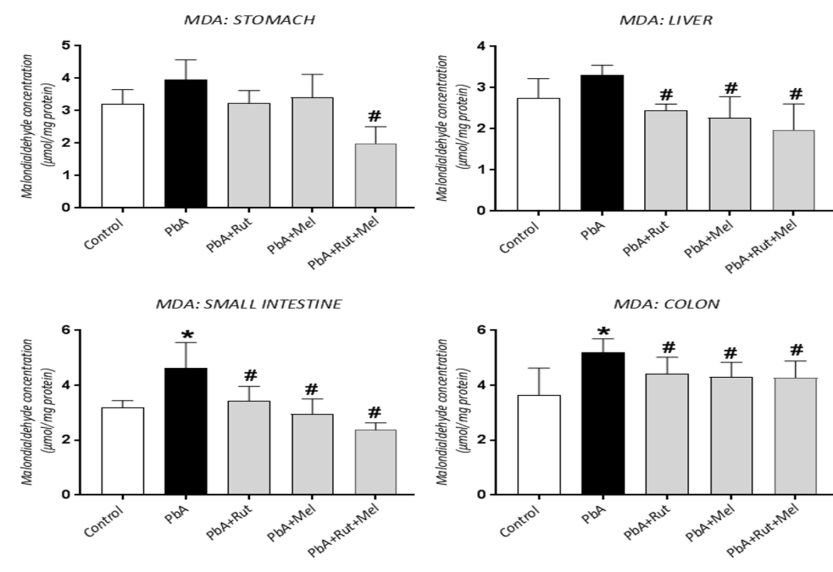
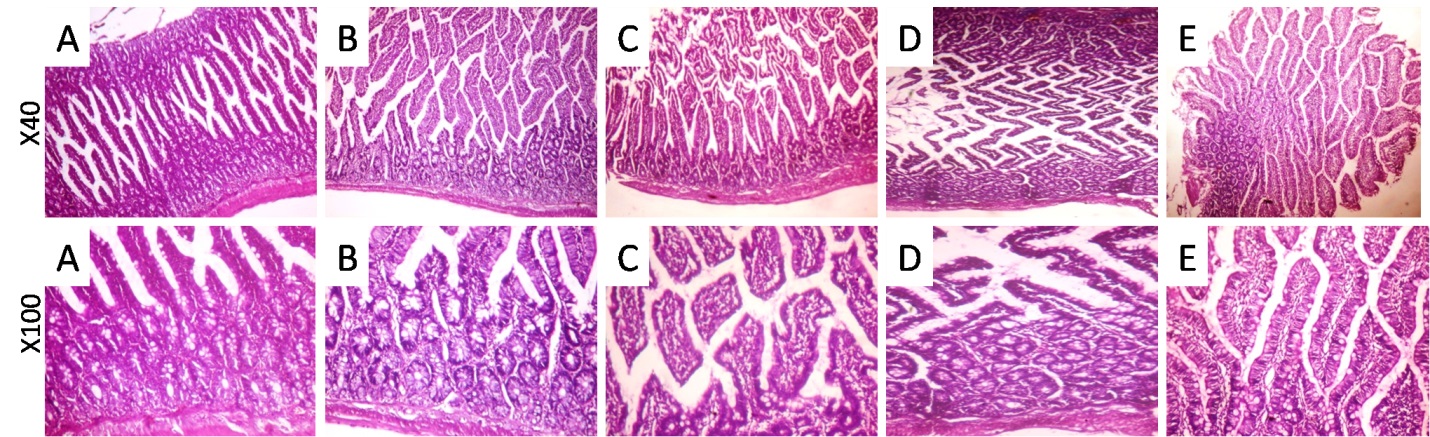
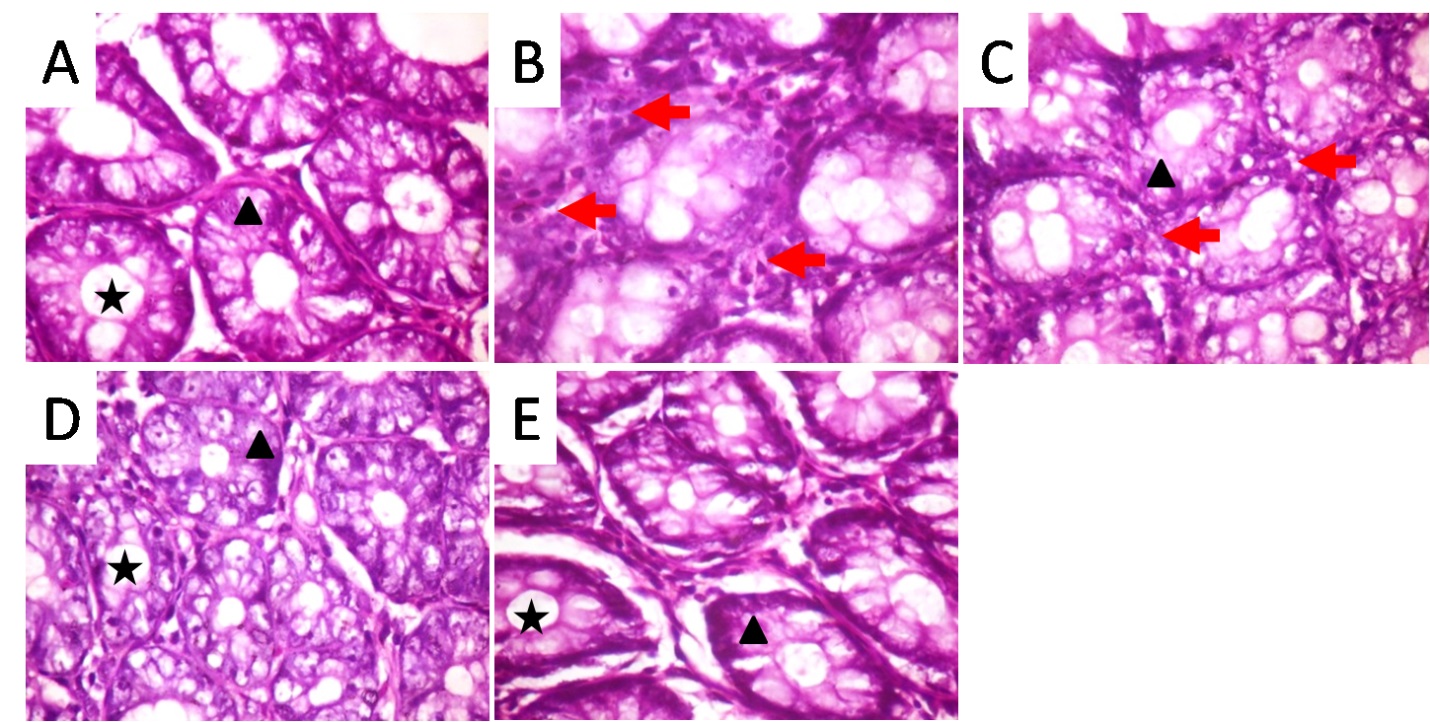
1. Luo L, Wang B, Jiang J, Fitzgerald M, Huang Q, Yu Z, et al. Heavy Metal Contaminations in Herbal Medicines: Determination, Comprehensive Risk Assessments, and Solutions. Frontiers in pharmacology. 2020;11:595335. [
DOI:10.3389/fphar.2020.595335] [
PMID] [
PMCID]
2. Omobowale TO, Oyagbemi AA, Akinrinde AS, Saba AB, Daramola OT, Ogunpolu BS, et al. Failure of recovery from lead induced hepatoxicity and disruption of erythrocyte antioxidant defence system in Wistar rats. Environmental toxicology and pharmacology. 2014;37(3):1202-11. [
DOI:10.1016/j.etap.2014.03.002] [
PMID]
3. Nkwunonwo UC, Odika PO, Onyia NI. A Review of the Health Implications of Heavy Metals in Food Chain in Nigeria. TheScientificWorldJournal. 2020;2020:6594109. [
DOI:10.1155/2020/6594109] [
PMID] [
PMCID]
4. de Carvalho C, Caramujo MJ. The Various Roles of Fatty Acids. Molecules. 2018;23(10). [
DOI:10.3390/molecules23102583] [
PMID] [
PMCID]
5. Calder PC. Fatty acids and inflammation: the cutting edge between food and pharma. European journal of pharmacology. 2011;668 Suppl 1:S50-8. [
DOI:10.1016/j.ejphar.2011.05.085] [
PMID]
6. Tang H, Zhu X, Gong C, Liu H, Liu F. Protective effects and mechanisms of omega-3 polyunsaturated fatty acid on intestinal injury and macrophage polarization in peritoneal dialysis rats. Nephrology. 2019;24(10):1081-9. [
DOI:10.1111/nep.13587] [
PMID] [
PMCID]
7. Zhang CX, Shu CM, Zhang XY, Lin XT, Guan QH, Zhang F. Effect and mechanism of omega-3 polyunsaturated fatty acids on intestinal injury in rats with obstructive jaundice. Europe Rev Med Pharmacol Sci. 2021;25(19):6077-92.
8. Tan J, McKenzie C, Potamitis M, Thorburn AN, Mackay CR, Macia L. The role of short-chain fatty acids in health and disease. Advances in immunology. 2014;121:91-119. [
DOI:10.1016/B978-0-12-800100-4.00003-9] [
PMID]
9. Blaak EE, Canfora EE, Theis S, Frost G, Groen AK, Mithieux G, et al. Short chain fatty acids in human gut and metabolic health. Beneficial microbes. 2020;11(5):411-55. [
DOI:10.3920/BM2020.0057] [
PMID]
10. Nogal A, Valdes AM, Menni C. The role of short-chain fatty acids in the interplay between gut microbiota and diet in cardio-metabolic health. Gut microbes. 2021;13(1):1-24. [
DOI:10.1080/19490976.2021.1897212] [
PMID] [
PMCID]
11. Song EM, Byeon JS, Lee SM, Yoo HJ, Kim SJ, Lee SH, et al. Fecal Fatty Acid Profiling as a Potential New Screening Biomarker in Patients with Colorectal Cancer. Digestive diseases and sciences. 2018;63(5):1229-36. [
DOI:10.1007/s10620-018-4982-y] [
PMID]
12. Sitkin SI, Tkachenko EI, Vakhitov TY. Metabolic dysbiosis of the gut microbiota and its biomarkers. Exprim Clinic Gastroenterol. 2016;12(12):6-29. [
DOI:10.18786/2072-0505-2015-40-12-34]
13. Zhuang X, Li T, Li M, Huang S, Qiu Y, Feng R, et al. Systematic Review and Meta-analysis: Short-Chain Fatty Acid Characterization in Patients With Inflammatory Bowel Disease. Inflammatory bowel diseases. 2019;25(11):1751-63. [
DOI:10.1093/ibd/izz188] [
PMID]
14. De Preter V, Machiels K, Joossens M, Arijs I, Matthys C, Vermeire S, et al. Faecal metabolite profiling identifies medium-chain fatty acids as discriminating compounds in IBD. Gut. 2015;64(3):447-58. [
DOI:10.1136/gutjnl-2013-306423] [
PMID]
15. Weir TL, Manter DK, Sheflin AM, Barnett BA, Heuberger AL, Ryan EP. Stool microbiome and metabolome differences between colorectal cancer patients and healthy adults. PloS one. 2013;8(8):e70803. [
DOI:10.1371/journal.pone.0070803] [
PMID] [
PMCID]
16. Wiktorowska-Owczarek A, Berezinska M, Nowak JZ. PUFAs: Structures, Metabolism and Functions. Advances in clinical and experimental medicine : official organ Wroclaw Medical University. 2015;24(6):931-41. [
DOI:10.17219/acem/31243] [
PMID]
17. Ravindran V, Tancharoenrat P, Zaefarian F, Ravindran G. Fats in poultry nutrition: Digestive physiology and factors influencing their utilization. Animal Feed Sci Technol. 2016;213:1-21. [
DOI:10.1016/j.anifeedsci.2016.01.012]
18. Han L, Wang H, Li L, Li X, Ge J, Reiter RJ, et al. Melatonin protects against maternal obesity-associated oxidative stress and meiotic defects in oocytes via the SIRT3-SOD2-dependent pathway. Journal of pineal research. 2017;63(3). [
DOI:10.1111/jpi.12431] [
PMID]
19. Liu Z, Gan L, Luo D, Sun C. Melatonin promotes circadian rhythm-induced proliferation through Clock/histone deacetylase 3/c-Myc interaction in mouse adipose tissue. Journal of pineal research. 2017;62(4). [
DOI:10.1111/jpi.12383] [
PMID]
20. Ganeshpurkar A, Saluja AK. The Pharmacological Potential of Rutin. Saudi pharmaceutical journal : SPJ : the official publication of the Saudi Pharmaceutical Society. 2017;25(2):149-64. [
DOI:10.1016/j.jsps.2016.04.025] [
PMID] [
PMCID]
21. Yong DOC, Saker SR, Chellappan DK, Madheswaran T, Panneerselvam J, Choudhury H, et al. Molecular and Immunological Mechanisms Underlying the Various Pharmacological Properties of the Potent Bioflavonoid, Rutin. Endocrine, metabolic & immune disorders drug targets. 2020;20(10):1590-6. [
DOI:10.2174/1871530320666200503053846] [
PMID]
22. Abdel-Raheem IT. Gastroprotective effect of rutin against indomethacin-induced ulcers in rats. Basic & clinical pharmacology & toxicology. 2010;107(3):742-50. [
DOI:10.1111/j.1742-7843.2010.00568.x] [
PMID]
23. Olaleye MT, Akinmoladun AC. Comparative gastroprotective effect of post-treatment with low doses of rutin and cimetidine in rats. Fundamental & clinical pharmacology. 2013;27(2):138-45. [
DOI:10.1111/j.1472-8206.2011.00972.x] [
PMID]
24. Chen CQ, Fichna J, Bashashati M, Li YY, Storr M. Distribution, function and physiological role of melatonin in the lower gut. World journal of gastroenterology. 2011;17(34):3888-98. [
DOI:10.3748/wjg.v17.i34.3888] [
PMID] [
PMCID]
25. Bhattacharya S, Patel KK, Dehari D, Agrawal AK, Singh S. Melatonin and its ubiquitous anticancer effects. Molecular and cellular biochemistry. 2019;462(1-2):133-55. [
DOI:10.1007/s11010-019-03617-5] [
PMID]
26. Kvetnoy IM, Ingel IE, Kvetnaia TV, Malinovskaya NK, Rapoport SI, Raikhlin NT. Gastrointestinal melatonin: cellular identification and biological role. Neuro Endocrinol Letter. 2002;23:121-32.
27. Bubenik GA. Gastrointestinal melatonin: localization, function, and clinical relevance. Digestive diseases and sciences. 2002;47(10):2336-48. [
DOI:10.1023/A:1020107915919] [
PMID]
28. Bothorel B, Barassin S, Saboureau M, Perreau S, Vivien-Roels B, Malan A, et al. In the rat, exogenous melatonin increases the amplitude of pineal melatonin secretion by a direct action on the circadian clock. The European journal of neuroscience. 2002;16(6):1090-8. [
DOI:10.1046/j.1460-9568.2002.02176.x] [
PMID]
29. Vyskocil A, Fiala Z, Tejnorová I, Tusl M. Stress reaction in developing rats exposed to 1% lead acetate. Sb Ved Pr Lek Fak Karlovy Univerzity Hradci Kralove. 1991;34(3):287-95.
30. Vyskocil A, Smejkalová J, Lacinová V. Dose-related stress reaction in male rats chronically exposed to lead acetate. Sb Ved Pr Lek Fak Karlovy Univerzity Hradci Kralove. 1991;34(5):393-401.
31. Mugahi MN, Heidari Z, Sagheb HM, Barbarestani M. Effects of Chronic Lead Acetate Intoxication on Blood Indices of Male Adult Rat. DARU J Pharmaceut Sci. 2003;11(4):147-51.
32. Ogeturk M, Kus I, Kavakli A, Zararsiz I, Ilhan N, Sarsilmaz M. Effects of melatonin on carbon tetrachloride-induced changes in rat serum. Journal of physiology and biochemistry. 2004;60(3):205-10. [
DOI:10.1007/BF03167030] [
PMID]
33. Demirtas CY, Pasaoglu OT, Bircan FS, Kantar S, Turkozkan N. The investigation of melatonin effect on liver antioxidant and oxidant levels in fructose-mediated metabolic syndrome model European Reviews for Medical and Pharmacological Sciences. 2015;19(10):1915-21.
34. Caglayan C, Kandemir FM, Yildirim S, Kucukler S, Eser G. Rutin protects mercuric chloride-induced nephrotoxicity via targeting of aquaporin 1 level, oxidative stress, apoptosis and inflammation in rats. Journal of trace elements in medicine and biology : organ of the Society for Minerals and Trace Elements. 2019;54:69-78. [
DOI:10.1016/j.jtemb.2019.04.007] [
PMID]
35. Kandemir FM, Caglayan C, Aksu EH, Yildirim S, Kucukler S, Gur C. Protective effect of rutin on mercuric chloride-induced reproductive damage in male rats. Andrologia. 2020;52(3):e13524. [
DOI:10.1111/and.13524]
36. Wolff SF. Ferrous ion oxidation in the presence of ferric ion indicator xylenol orange for measurement of hydrogen peroxides. Method Enzymol. 1994;233(2):182-9. [
DOI:10.1016/S0076-6879(94)33021-2]
37. Varshney R, Kale RK. Effects of calmodulin antagonists on radiation-induced lipid peroxidation in microsomes. International journal of radiation biology. 1990;58(5):733-43. [
DOI:10.1080/09553009014552121] [
PMID]
38. Ellman GL. Tissue sulfhydryl groups. Archives of biochemistry and biophysics. 1959;82(1):70-7. [
DOI:10.1016/0003-9861(59)90090-6] [
PMID]
39. Jollow DJ, Mitchell JR, Zampaglione N, Gillette JR. Bromobenzene-induced liver necrosis. Protective role of glutathione and evidence for 3,4-bromobenzene oxide as the hepatotoxic metabolite. Pharmacology. 1974;11(3):151-69. [
DOI:10.1159/000136485] [
PMID]
40. Habig WH, Pabst MJ, Jakoby WB. Glutathione-S-transferases. The first enzymatic step in mercapturic acid formation. J Biol Chemist. 1974;25:7130-9. [
DOI:10.1016/S0021-9258(19)42083-8]
41. Misra HP, Fridovich I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. The Journal of biological chemistry. 1972;247(10):3170-5. [
DOI:10.1016/S0021-9258(19)45228-9]
42. Oyagbemi AA, Omobowale TO, Akinrinde AS, Saba AB, Ogunpolu BS, Daramola O. Lack of reversal of oxidative damage in renal tissues of lead acetate-treated rats. Environmental toxicology. 2015;30(11):1235-43. [
DOI:10.1002/tox.21994] [
PMID]
43. Scortichini S, Boarelli MC, Silvi S, Fiorini D. Development and validation of a GC-FID method for the analysis of short chain fatty acids in rat and human faeces and in fermentation fluids. Journal of chromatography B, Analytical technologies in the biomedical and life sciences. 2020;1143:121972. [
DOI:10.1016/j.jchromb.2020.121972] [
PMID]
44. Akinrinde A, Adigun K, Mustapha A. Cobalt-induced neuro-behavioural alterations are accompanied by profound Purkinje cell and gut-associated responses in rats. Environ Anal Health Toxicol. 2023;38(2):e2023010-0. [
DOI:10.5620/eaht.2023010]
45. Crespo S, Nonnotte G, Colin DA, Leray C, Aubree A. Morphological and functional alterations induced in trout intestine by dietary cadmium and lead. J Fish Biol. 2010;28:69-80. [
DOI:10.1111/j.1095-8649.1986.tb05143.x]
46. Cheng D, Li H, Zhou J, Wang S. Chlorogenic acid relieves lead-induced cognitive impairments and hepato-renal damage via regulating the dysbiosis of the gut microbiota in mice. Food & function. 2019;10(2):681-90. [
DOI:10.1039/C8FO01755G] [
PMID]
47. Zhai Q, Qu D, Feng S, Yu Y, Yu L, Tian F, et al. Oral Supplementation of Lead-Intolerant Intestinal Microbes Protects Against Lead (Pb) Toxicity in Mice. Frontiers in microbiology. 2019;10:3161. [
DOI:10.3389/fmicb.2019.03161] [
PMID] [
PMCID]
48. Marion-Letellier R, Savoye G, Beck PL, Panaccione R, Ghosh S. Polyunsaturated fatty acids in inflammatory bowel diseases: a reappraisal of effects and therapeutic approaches. Inflammatory bowel diseases. 2013;19(3):650-61. [
DOI:10.1097/MIB.0b013e3182810122] [
PMID]
49. Gobbetti T, Dalli J, Colas RA, Federici Canova D, Aursnes M, Bonnet D, et al. Protectin D1(n-3 DPA) and resolvin D5(n-3 DPA) are effectors of intestinal protection. Proceedings of the National Academy of Sciences of the United States of America. 2017;114(15):3963-8. [
DOI:10.1073/pnas.1617290114] [
PMID] [
PMCID]
50. Zhu H, Wang H, Wang S, Tu Z, Zhang L, Wang X, et al. Flaxseed Oil Attenuates Intestinal Damage and Inflammation by Regulating Necroptosis and TLR4/NOD Signaling Pathways Following Lipopolysaccharide Challenge in a Piglet Model. Molecular nutrition & food research. 2018;62(9):e1700814. [
DOI:10.1002/mnfr.201700814] [
PMID]
51. Lauridsen C. Lipid nutrition and gut health of pigs. J Animal Sci. 2019;97(Supplement 2):28-54. [
DOI:10.1093/jas/skz122.051] [
PMCID]
52. Wang Y, Chen Y, Zhang X, Lu Y, Chen H. New insights in intestinal oxidative stress damage and the health intervention effects of nutrients: A review. J Funct Food. 2020;75:104248. [
DOI:10.1016/j.jff.2020.104248]
53. Rosas CE, Correa LB, Henriques MG. Role of Neutrophils in Disease Pathogenesis. London, UK: Intech Open Limited. Neutrophils in rheumatoid arthritis: a target for discovering new therapies based on natural products.2017. 89-118 p. [
DOI:10.5772/intechopen.68617]
54. Rana SV, Sharma S, Prasad KK, Sinha SK, Singh K. Indian Journal of Medical Research. 2014;139(4):568-71.
55. Yu L, Yu Y, Xiao Y, Tian F, Narbad A, Zhai Q. Lead-induced gut injuries and the dietary protective strategies: A review. J Funct Food. 2021;83:104528. [
DOI:10.1016/j.jff.2021.104528]
56. Hoyles L, Wallace RJ. Gastrointestinal Tract: Intestinal Fatty Acid Metabolism and Implications for Health. Handbook Hydrocarbon Lipid Microbiol. 2010:3119-32. [
DOI:10.1007/978-3-540-77587-4_234]
57. Hassan FAM, Roushdy EM, Kishawy ATY, Zaglool AW, Tukur HA, Saadeldin IM. Growth Performance, Antioxidant Capacity, Lipid-Related Transcript Expression and the Economics of Broiler Chickens Fed Different Levels of Rutin. Animals : an open access journal from MDPI. 2018;9(1). [
DOI:10.3390/ani9010007] [
PMID] [
PMCID]
58. Yang Z, He Y, Wang H, Zhang Q. Protective effect of melatonin against chronic cadmium-induced hepatotoxicity by suppressing oxidative stress, inflammation, and apoptosis in mice. Ecotoxicology and environmental safety. 2021;228:112947. [
DOI:10.1016/j.ecoenv.2021.112947] [
PMID]
59. Ghosh D, Dey M, Ghosh A, Chattopadhyay A, Bandyopadhyay D. Melatonin protects against lead acetate-induced changes in blood corpuscles and lipid profile of male Wistar rats. J Pharmacy Res. 2014;8(3):336-42.
60. Hernandez-Plata E, Quiroz-Compean F, Ramirez-Garcia G, Barrientos EY, Rodriguez-Morales NM, Flores A, et al. Melatonin reduces lead levels in blood, brain and bone and increases lead excretion in rats subjected to subacute lead treatment. Toxicology letters. 2015;233(2):78-83. [
DOI:10.1016/j.toxlet.2015.01.009] [
PMID]
61. Ilesanmi OB, Adeogun EF, Odewale TT, Chikere B. Lead exposure-induced changes in hematology and biomarkers of hepatic injury: protective role of TrevoTM supplement. Environ Anal Health Toxicol. 2022;37(2):e2022007-0. [
DOI:10.5620/eaht.2022007] [
PMID] [
PMCID]