Introduction
A vital component of general health is male reproductive health, and worries about fertility have increased in recent years [1]. Any effect of chemicals that would interfere with reproductive ability or potential is referred to as reproductive toxicity, which has an impact on lactation and the development of the offspring [2]. The reproductive cycle, a complex biological function, can be altered by toxic exogenous substances, making it susceptible to reproductive toxicity. Household products, such as mosquito coils, insecticides, incense, and aspirin, can induce reproductive toxicity, potentially leading to infertility issues in individuals [3-6]. Common vegetables, including kale and red onions, are rich in quercetin, a natural product and dietary supplement that has anti-inflammatory and antioxidant properties. The anti-inflammatory, antipyretic, and analgesic properties of aspirin, a non-steroidal anti-inflammatory medication, pose many benefits in this regard [7]. According to numerous studies, aspirin has drawn a lot of interest as a cardio-protective agent in addition to being an analgesic [8].
The main reproductive function of male fertility, sperm generation, is impacted by a number of chemicals, radiation, environmental conditions, and therapeutic and non-therapeutic agents [9]. As stated in a recent report by the World Health Organization, infertility will affect a significant proportion of people at some point in their lives [10]. Spermatozoa exposure to oxidative stress is a major contributing factor to male infertility [11], with sperm DNA fragmentation being a key factor [12]. A high level of 8-OHdG oxidative DNA damage is linked to male infertility and aberrant spermatozoa [13]. In light of this, the present research attempted to examine the possible influence of quercetin on male fertility, with a particular emphasis on how it affects testicular tissues. Both immunohistochemical and morphological investigations will be used in the evaluation, with a focus on sperm parameters and histomorphology.
Materials and Methods
Animals and housing conditions
In this study, 35 adult male Wistar rats weighing 150–200 grams were kept in plastic cages in the animal house of the Department of Anatomy, Faculty of Basic Medical Sciences, Ekiti State University, Ado-Ekiti, Nigeria, under conventional laboratory conditions. In accordance with the Guidelines of the National Institute of Health for the Care and Use of Laboratory Animals, they were given pelleted meal and unlimited water [14].
Drugs and chemicals
Quercetin (Sigma–Aldrich, Milan, Italy) and an Aspirin tablet 50 mg (Juli Pharmaceutical Ltd, Ekiti, Nigeria) were purchased. All other chemicals used in this study were obtained commercially, and they were of analytical grade as appropriate. The powdered form of Quercetin was dissolved in distilled water and administered orally to rats as a suspension in distilled water for eight weeks with the use of an oral cannula.
Experimental design
Rats were divided into seven experimental groups at random (n=5 per group). The groups consisted of two control groups, each of which received 9.30 mL/kg body weight of normal saline and olive oil; the Quercetin group, which received 30 mg/kg body weight of Quercetin alone; the Quercetin group, which received 60 mg/kg body weight of Quercetin alone; the Aspirin group, which received 50 mg/kg body weight of Aspirin alone; the Aspirin + Quercetin group, which received 30 mg/kg body weight of Quercetin and 50 mg/kg body weight of Aspirin; and the Aspirin + Quercetin group, which received 60 mg/kg body weight of Quercetin and 50 mg/kg body weight. Body weights of the rats were measured both before and after each week of the experiment.
Animal sacrifice and tissue collection
After 24 h of the last treatment, animals were euthanized with 80 mg/kg ketamine, and the caudal part of their epididymis and testis were excised and weighed, followed by sperm analysis. The entire body was fixed with 4% paraformaldehyde.
Sperm parameter analysis
Sperm count, motility, and morphology were evaluated using Computer Assisted Sperm Analysis (Microptic S.L., Carrer de la Ciutat de Granada, 131, 08018 Barcelona, Spain).
Histological analysis
Testis tissues were fixed in 10% formalin, dehydrated with ethanol, cleared with xylene, embedded in wax, cut, stained with hematoxylin and eosin, and microscopically analyzed.
Immuno-histochemical analysis
The sections were dewaxed, hydrated, and immersed in citrate buffer for 40 min. After cooling, they were washed with phosphate-buffered saline, and hydrogen peroxide was applied and incubated with the primary antibody. Biotinylated link and Streptavidin horseradish peroxidase reagent were applied. The 3,3'-Diaminobenzidine substrate chromogen solution was applied, and human colon adenocarcinoma was used as a positive control. Negative control slides were prepared using the antibody diluent [15]. Percentage of Proliferating Cell Nuclear Antigen (PCNA)-positive areas was estimated in all groups and digitized using an Olympus® digital camera (OM Digital Solutions Corporation, 49-3 Takakura-machi, Hachioji-shi, Tokyo 192-0033, Japan) and an Olympus® microscope at 40 × magnification.
Ethical approval
The Ethics and Research Committee of the College of Medicine at Ekiti State University, Ado-Ekiti, granted ethical permission for this investigation (ERCANA/2023/12/001). The National Institutes of Health's Guidelines for the Care and Use of Laboratory Animals were followed in the methods and procedures of the experiment [14].
Statistical analysis
Data was tested using analysis of variance to determine statistical significance at p < 0.05, and the findings were presented as mean ± standard error of the mean. GraphPad Prism 5 (GraphPad, San Diego, CA, USA) was used to statistically assess sperm analysis, motility, morphology, and quantitative data in order to find any correlations or differences. Statistical significance was set at p < 0.05.
Results
Table 1 summarizes the gross anatomical measurements across the experimental groups. The control, olive oil, and quercetin-only groups all showed comparable increases in body weight throughout the study period, reflecting normal growth. Rats that received aspirin displayed the smallest increase in body weight and the lowest testicular weight and volume. In the groups receiving both aspirin and quercetin, testicular weight and volume were higher than in the aspirin group, with values approaching those of the untreated controls.
Table 1. Gross anatomical analysis (body weight, testicular weight and volume).
| Groups |
Initial body weight (g) |
Final body weight (g) |
Body weight diff (g) |
Testis weight (g) |
Testis volume (ml) |
Testis weight/body weight ratio |
| Control |
210 ± 3.5 |
233 ± 3.3 |
23 ± 0.2 |
1.313 ± 0.96 |
1.100 ± 0.23 |
0.004 |
| Olive oil |
208 ± 3.2 |
230 ± 3.6 |
22 ± 0.4 |
1.207 ± 0.31 |
0.966 ± 0.03 |
0.004 |
| Que 30 mg |
207± 2.3 |
230 ± 0.9 |
23 ± 1.4 |
1.310 ± 0.15 |
1.067 ± 0.03 |
0.005 |
| Que 60 mg |
209 ± 4.3 |
229 ± 4.1 |
20 ± 0.2 |
1.273 ± 0.20 |
1.000 ± 0.05 |
0.005 |
| Asp 50 mg |
202 ± 4.1 |
210 ± 5.3 |
8 ± 1.2 |
0.953 ± 0.02 |
1.000 ± 0.00 |
0.002 |
| Que 30 mg + Asp 50 mg |
201 ± 1.0 |
218 ± 1.7 |
17 ± 0.7 |
1.243 ± 0.02 |
1.033 ± 0.03 |
0.003 |
| Que 60 mg + Asp 50 mg |
206 ± 3.2 |
219 ± 3.8 |
13 ± 0.6 |
1.234 ± 0.03 |
1.067 ± 0.03 |
0.004 |
Que: Quercetin, Asp: Aspirin. Values are reported in mean ± standard error of the mean.
Figures 1 to 3 illustrate the quantitative sperm parameters. In the control, olive oil, and quercetin-only groups, sperm count, motility, and morphology remained within a similar range. Aspirin administration was associated with the lowest sperm concentration, reduced progressive motility, and a higher proportion of abnormal spermatozoa. In both co-treatment groups, sperm count increased relative to aspirin alone. Motility measurements also showed a clear upward shift, and a greater percentage of spermatozoa retained normal morphology. The improvement was observed across all three parameters, indicating that the presence of quercetin altered the pattern observed in aspirin-only animals without restoring values fully to control levels. The visual representation in Figures 1 to 3 highlights these group differences, with the aspirin group consistently occupying the lowest position across all endpoints and the co-treated groups positioned between the aspirin and quercetin-only groups.
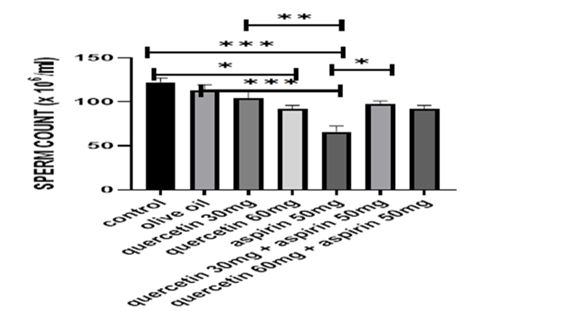
Figure 1. Sperm count (x 106/mL) differences among different groups. Data are presented as mean ± standard error of the mean (n = 5). Statistical analysis was performed using one-way analysis of variance. *p < 0.05, **p < 0.01, ***p < 0.001.
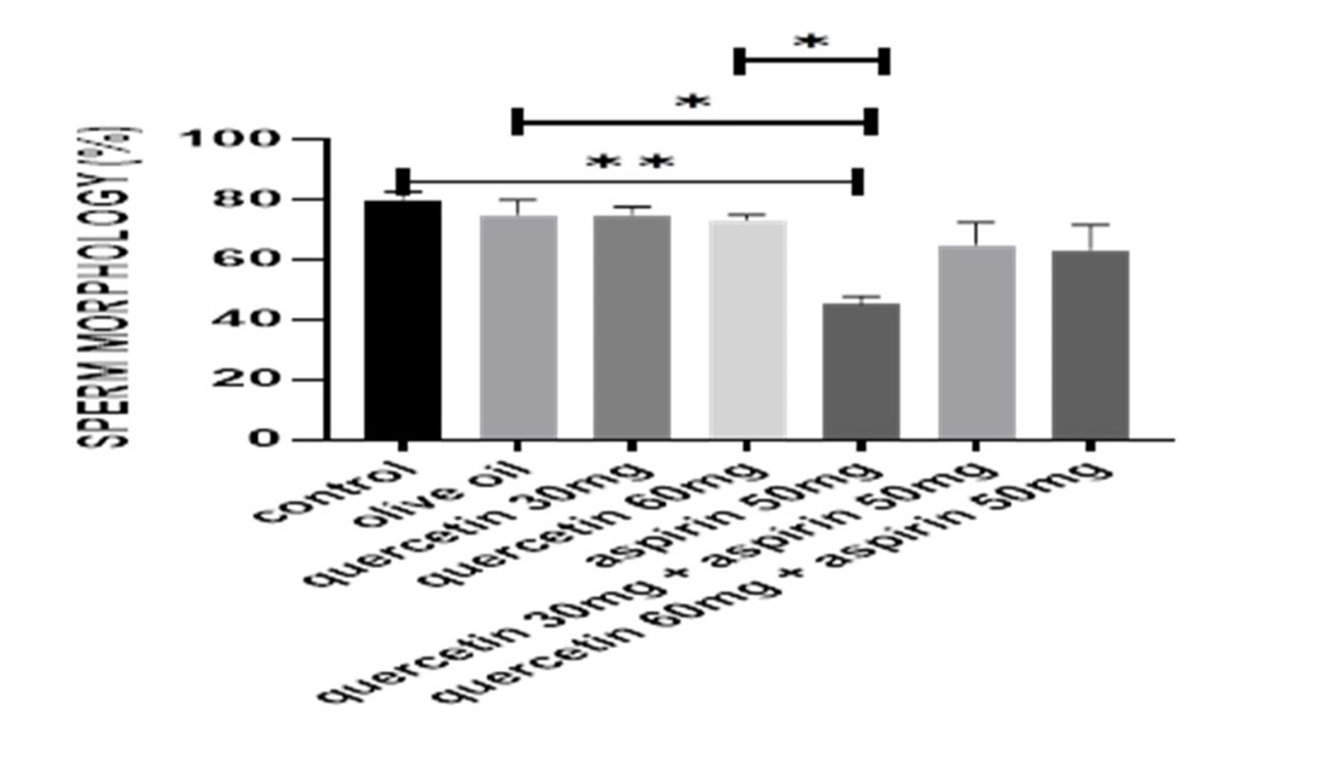
Figure 2. Sperm morphology changes (%) of different groups. Data are presented as mean ± standard error of the mean (n = 5). Statistical analysis was performed using one-way analysis of variance. *p < 0.05, **p < 0.01, ***p < 0.001.
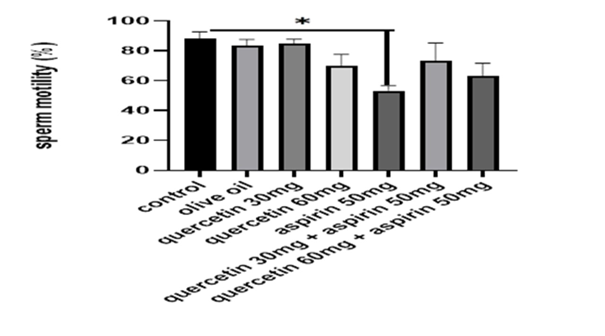
Figure 3. Sperm motility changes (%) of different groups. Data are presented as mean ± standard error of the mean (n = 5). Statistical analysis was performed using one-way analysis of variance. *p < 0.05, **p < 0.01, ***p < 0.001.
Histological findings are shown in Figure 4. Testicular sections from the control, olive oil, and quercetin-only groups demonstrated intact seminiferous tubules with well-defined germinal layers and lumens, as well as the presence of maturing germ cells. Sections from aspirin-treated rats displayed a reduced number of germ cell layers, wider luminal spaces, and disrupted cellular arrangement. The groups receiving quercetin alongside aspirin exhibited seminiferous tubules with more ordered cell layers and less luminal widening than the aspirin-only group. Although some alterations remained visible, the overall structure appeared more preserved in the co-treated groups.
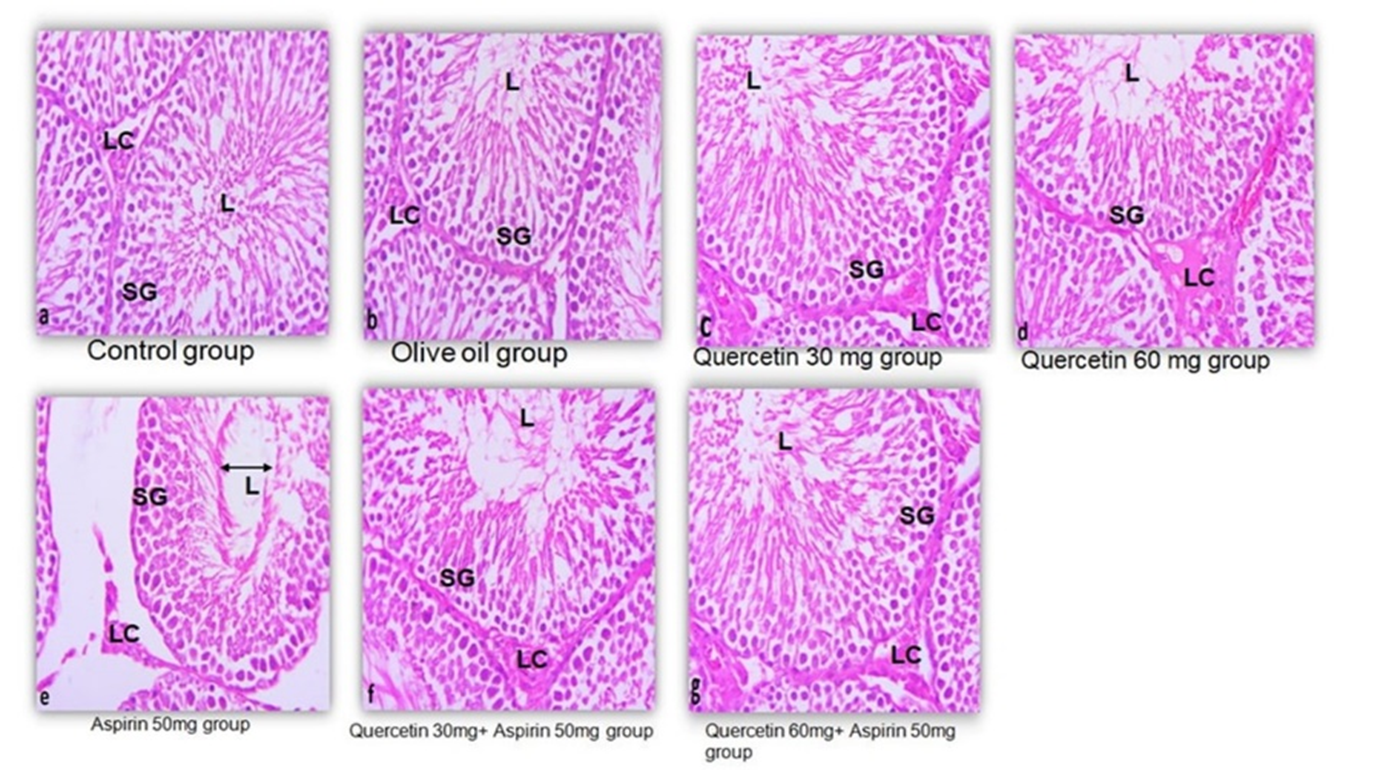
Figure 4. Histoarchitecture of control groups showing normal testicular architecture with an orderly arrangement of normal seminiferous tubules and an adequate number of sperm cells. a) control group; b) olive oil group, while treatment with quercetin for 30 days resulted in well-organized structure in testis histology; c-d) the primary and secondary spermatogonia, spermatocytes, and Leydig’s cells exhibited significant reduction (p ≤ 0.001 for all) following administration of aspirin; and e) sections of quercetin + aspirin, compared to vehicle control were mildly significantly reduced; f-g) Leydig cells (LC), Lumen of seminiferous epithelium (L), spermatogonia (SG).
The PCNA immunostaining patterns are presented in Figure 5. Strong nuclear staining was evident in the basal germ cells of the control, olive oil, and quercetin-only groups. Aspirin-treated sections showed reduced staining intensity and fewer PCNA-positive cells. In both co-treated groups, the number of positively stained germ cells exceeded that of the aspirin-only group. The group receiving 60 mg/kg quercetin showed the most extensive PCNA reactivity; however, the pattern did not fully replicate that of the control groups.
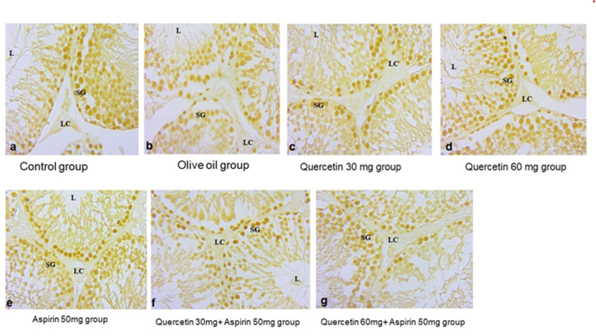 Figure 5. Photomicrographs of sections in testicular tissue (proliferating cell nuclear antigen immunostaining) showing: a) control group with positive immunostaining (brown nuclear reaction) in most of the basal germ cells in the seminiferous tubules; b) olive oil; c) quercetin 30 mg; and d) quercetin 60 mg showing narrow interstitial tissue containing Leydig cells (LC), Lumen of seminiferous epithelium (L), spermatogonia (SG) as shown in control group; (e) aspirin-treated group with few immuno-positive germ cells; f) aspirin 50 mg + quercetin 30 mg treated group with more positive basal germ cells similar to g) aspirin 50 mg + quercetin 60 mg, which shows more other mild positive germ cells, compared to only aspirin-treated groups.
Figure 5. Photomicrographs of sections in testicular tissue (proliferating cell nuclear antigen immunostaining) showing: a) control group with positive immunostaining (brown nuclear reaction) in most of the basal germ cells in the seminiferous tubules; b) olive oil; c) quercetin 30 mg; and d) quercetin 60 mg showing narrow interstitial tissue containing Leydig cells (LC), Lumen of seminiferous epithelium (L), spermatogonia (SG) as shown in control group; (e) aspirin-treated group with few immuno-positive germ cells; f) aspirin 50 mg + quercetin 30 mg treated group with more positive basal germ cells similar to g) aspirin 50 mg + quercetin 60 mg, which shows more other mild positive germ cells, compared to only aspirin-treated groups.
Discussion
During this study, there was no mortality observed among the study groups; however, rats treated with aspirin demonstrated a notable decrease in testicular and body weights, compared to the quercetin-only and control groups, indicating potential reproductive toxicity (Table 1). In contrast, the quercetin-only and olive oil groups maintained comparable weights to the vehicle-treated controls, suggesting the non-toxic effect of quercetin and the vehicle. Sperm analysis (Figures 1–3) revealed marked reductions in sperm motility, count, and usual morphology in the aspirin-only group, corroborating the detrimental impact of aspirin on spermatogenesis. These parameters were significantly improved in the co-treated groups (aspirin + quercetin at both 30 mg/kg and 60 mg/kg), demonstrating the potential of quercetin in mitigating aspirin-induced reproductive damage (p < 0.05). Preservation of sperm quality in these groups suggests that quercetin supports spermatogenic integrity and functionality even under oxidative stress conditions induced by aspirin.
Histological assessments provided further insights. Accordingly, in the control and quercetin-only groups (Figure 4a–b), testicular architecture was intact, with well-organized seminiferous tubules, stratified germinal epithelium, and evident mature spermatozoa. These findings confirmed normal spermatogenesis and tissue homeostasis. In contrast, aspirin-treated rats exhibited severe disruptions in testicular structure, including sloughing of the basal lamina, widened tubular lumens, and disorganized spermatogenic cells (Figure 4e), aligning with prior studies that highlighted the capacity of aspirin to compromise testicular tissue integrity [16, 17]. Notably, co-administration of quercetin with aspirin (Figures 4f–g) showed partial preservation of the seminiferous tubules and interstitial tissues, with improved cellular organization and germ cell presence. Although some features remained mildly affected, the degree of restoration was substantial, especially at the higher quercetin dose, suggesting dose-dependent protection. These histological findings supported previous evidence that quercetin can scavenge free radicals and limit glutathione depletion, thus preserving testicular architecture and function [18, 19].
Histological analysis further supported these findings by quantifying cellular integrity. In aspirin-treated rats, there was a significant reduction in spermatogonia and Leydig cells, all crucial for spermatogenesis and steroidogenesis (Figure 4e). Restoration of these cells in the quercetin co-treatment groups (Figure 4f–g) aligned with the known role of quercetin in maintaining cellular proliferation and reducing apoptotic damage [20]. The observed reduction in testicular weight in the aspirin-treated groups underscored the relationship between testis size and reproductive capacity. Reduced testicular weight is associated with decreased Sertoli cell numbers, daily generation of sperm, and testosterone yield [21, 22]. Interference of aspirin with these processes may be ascribed to its disruption of the blood-testis barrier and alteration of the microenvironment of seminiferous tubules [17]. Additionally, the shrinkage of seminiferous tubules and interstitial vacuolization seen histologically in these groups indicate compromised spermatogenic function, consistent with findings from earlier studies [23].
Functional effects of these morphological changes were further evidenced by PCNA immunostaining reaction (Figure 5). The PCNA, a marker of cellular growth, was robustly expressed in the spermatogonia of the control and quercetin-only groups, indicating active spermatogenesis. However, aspirin administration significantly reduced PCNA-positive cells, suggesting impaired cellular proliferation and increased apoptosis. While co-treatment with quercetin moderately improved PCNA reactivity, it remained lower than the control, indicating partial but meaningful recovery of proliferative capacity. These findings support previous reports highlighting the role of quercetin in modulating apoptotic pathways and enhancing cellular regeneration in testicular tissue [20, 24].
Conclusions
This study demonstrated that aspirin induces significant testicular toxicity, evidenced by reduced sperm quality, disrupted histological architecture, and diminished cellular proliferation. Quercetin, particularly at higher doses, effectively mitigated these adverse effects, restoring testicular structure and function. These findings highlight the potential of quercetin as a protective agent against drug-induced reproductive toxicity and support its therapeutic relevance in preserving male fertility.
Data Access and Responsibility
The authors confirm that this article contains original
work and accept full responsibility for its content.
Ethical Considerations
The Ethics and Research Committee of the College of Medicine at Ekiti State University, Ado-Ekiti, Nigeria, granted ethical permission for this investigation (ERCANA/2023/12/00l). Guidelines of the National Institutes of Health for the Care and Use of Laboratory Animals were followed in the methods and procedures of the experiment.
Authors' Contributions
A. M. A., and E. E. A. conceived and designed the study. All authors participated in the execution of the studies and the data collection. O. O., C. O. F., and A. F. conducted data analysis and wrote the manuscript draft. All authors reviewed and edited the final draft.
Conflict of Interests
The authors declare that there is no conflict of interest.
Funding
The authors received no financial support for the
research and publication of this article.
References
- Agarwal A, Baskaran S, Parekh N, Cho CL, Henkel R, Vij S, et al. Male infertility. Lancet. 2021;397(10271):319-33. [DOI:10.1016/S0140-6736(20)32667-2] [PMID:33308486]
- CHEMICALS LO. Globally harmonized system of classification and labelling of chemicals (GHS). 2002;1-37. [LINK]
- Akingbade AM, Saalu LC, Oyebanji OO, Oyeniran DA, Akande OO, Akunna GG. Rhodinol-based Incense Testiculotoxicity in Albino Rats: testicular histology, spermatogenic and biochemical evaluations. J Pharmacol Toxicol. 2014;9(2):68-81. [DOI:10.3923/jpt.2014.68.81]
- Akingbade AM, Ojewale AO, Idhirhi A, Olasehinde OR, Ibitoye OB, Aladeyelu SO. Testiculotoxicity activities of isopropyl alcohol-based air freshener on the testis of adult Wistar rats. J Mol Pathophysiol. 2017;6(2):17-23.[ DOI:10.5455/jmp.20170415095238]
- Akunna GG, Obikili EN, Anyawu GE, Esom EA. Evidences for spermatozoa toxicity and oxidative damage of cadmium exposure in rats. J Pharmacol Toxicol. 2017;12(1):50-6. [DOI:10.3923/jpt.2017.50.56 ]
- Akingbade MA, Adegoke-Kehinde O, Faeji CO, Fasoro A. Modulatory effect of quercetin on aspirin-induced hepatoxicity in Wistar rats. Indian J Physiol Pharmacol. 2025;69(1):34-39. [DOI:10.25259/IJPP_73_2024 ]
- Van Der Woude FJ, Heinemann LAJ, Graf H, Lewis M, Moehner S, Assmann A, et al. Analgesics use and ESRD in younger age: a case-control study. BMC Nephrol. 2007;8(1):15. [DOI:10.1186/1471-2369-8-15] [PMID: 18053232]
- Corman SL, Fedutes BA, Ansani NT. Impact of nonsteroidal antiinflammatory drugs on the cardioprotective effects of Aspirin. Ann Pharmacother. 2005;39(6):1073-9. [DOI:10.1345/aph.1E514] [PMID: 15870140]
- Meng Q, Ren A, Zhang L, Liu J, Li Z, Yang Y, et al. Incidence of infertility and risk factors of impaired fecundity among newly married couples in a Chinese population. Reprod Biomed Online. 2015;30(1):92-100.[DOI:10.1016/j.rbmo.2014.10.002] [PMID:25456165]
- Babakhanzadeh E, Nazari M, Ghasemifar S, Khodadadian A. Some of the factors involved in male infertility: a prospective review. Int J Gen Med. 2020;13:29-41. [DOI:10.2147/IJGM.S241099] [PMID:32104049]
- Aitken RJ, De Iuliis GN, Gibb Z, Baker MA. The simmet lecture: new horizons on an old landscape – oxidative stress, DNA damage and apoptosis in the male germ line. Reprod Domest Anim. 2012;47(4):7-14. [DOI:10.1111/j.1439-0531.2012.02049.x] [PMID: 22827344]
- Wright C, Milne S, Leeson H. Sperm DNA damage caused by oxidative stress: modifiable clinical, lifestyle and nutritional factors in male infertility. Reprod Biomed Online. 2014;28(6):684-703. [DOI:10.1016/j.rbmo.2014.02.004] [PMID:24745838]
- Guz J, Gackowski D, Foksinski M, Rozalski R, Zarakowska E, Siomek A, et al. Comparison of oxidative stress/DNA damage in semen and blood of fertile and infertile men. PLoS One. 2013;8(7):e68490. [DOI:10.1371/journal.pone.0068490] [PMID: 23874641]
- National research council. Guide for the care and Use of laboratory animals. 2011;1-246. [LINK]
- Altay B, Çetinkalp Ş, Doǧanavşargil B, Hekimgil M, Semerci B. Streptozotocin-induced diabetic effects on spermatogenesis with proliferative cell nuclear antigen immunostaining of adult rat testis. Fertil Steril. 2003;80(2)828-31. [DOI:10.1016/S0015-0282(03)00984-1] [PMID:14505760]
- Vyas R, Kesari KK, Slama P, Roychoudhury S, Sisodia R. Differential activity of antioxidants in testicular tissues following administration of chlorophytum borivilianum in Gamma-Irradiated swiss Albino mice. Front Pharmacol. 2022;12:774444. [DOI: 10.3389/fphar.2021.774444] [PMID: 35111049]
- Oyedeji KO, Kpaduwa PC. Effect of Aspirin on reproductive function in female wistar rats. J Pharm Sci Res. 2021;13(8):448-51. [LINK]
- Galati G, Moridani MY, Chan TS, O’Brien PJ. Peroxidative metabolism of apigenin and naringenin versus luteolin and quercetin: glutathione oxidation and conjugation. Free Radic Biol Med. 2001;30(4):370-82. [DOI:10.1016/S0891-5849(00)00481-0] [PMID:11182292]
- Hämäläinen M, Nieminen R, Vuorela P, Heinonen M, Moilanen E. Anti-Inflammatory effects of flavonoids: genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-κB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-κB activation along with their inhibitory effect on iNOS expression and NO Production in activated macrophages. Mediators Inflamm. 2007;2007(1):45673. [DOI:10.1155/2007/45673] [PMID:18274639]
- Lesser S, Cermak R, Wolffram S. The fatty acid pattern of dietary fat influences the oral bioavailability of the flavonol quercetin in pigs. Br J Nutr. 2006;96(6):1047-52. [DOI:10.1017/BJN20061953] [PMID:17181879]
- Thompson EL, Murphy KG, Patterson M, Bewick GA, Stamp GWH, Curtis AE, et al. Chronic subcutaneous administration of kisspeptin-54 causes testicular degeneration in adult male rats. Am J Physiol Endocrinol Metab. 2006;291(5):1074-82. [DOI:10.1152/ajpendo.00040.2006] [PMID:16787965]
- Cooke PS, Walker WH. Male fertility in mice requires classical and nonclassical androgen signaling. Cell Rep. 2021;36(7):109557. [DOI:10.1016/j.celrep.2021.109557] [PMID: 34407397]
- Vyas A, Ram H, Purohit A, Jatwa R. Adverse effects of subchronic dose of Aspirin on reproductive profile of male rats. J Pharm. 2016;2016(1):6585430. [DOI:10.1155/2016/6585430] [PMID:27190691]
- Cavalcante MB, Costa Fda S, Barini R, Araujo Júnior E. Granulocyte colony-stimulating factor and reproductive medicine: A review. Iran J Reprod Med. 2015;13(4):195-202. [PMID:26131007]




 , Ekpe Ekpe Asuquo2
, Ekpe Ekpe Asuquo2 



